Host-Parasite Interactions between Birds and Feather Mites 
2 Department of Wildlife and Safari Management, Chinhoyi University of Technology, Bag 7724 Chinhoyi, Zimbabwe
 Author
Author  Correspondence author
Correspondence author
International Journal of Super Species Research, 2012, Vol. 2, No. 1 doi: 10.5376/ijssr.2012.02.0001
Received: 25 May, 2012 Accepted: 28 Jun., 2012 Published: 05 Jul., 2012
Tarakini and Bastian., 2012, Host-Parasite Interactions between Birds and Feather Mites, International Journal of Super Species Research, 2012, Vol.2, No.1 1-6 (doi: 10.5376/ijssr.2012.02.0001)
There is current uncertainty on whether feather mites are a cause or consequence of poor body condition in birds. We aimed at investigating the bird-mite relationships and elucidating the functional significance of feather mites on birds found on the Urra Field station, Sorbas, in Almeria province, south-east Spain. We captured birds using mist nest and assessed birds for body condition (weight, fat and pectoral muscles), mite distribution on the wings and tested for diurnal changes in mite abundance. The Kruskal-Wallis test was used to test for differences in mite abundance across species, sex, age and to test for differences in mite distribution on wings across species while the Fisher`s exact test was used to test for diurnal mite abundance. There were no significant differences in mite abundance between males and females in blackcaps, house sparrows, Sardinian and Willow warblers. There were significant differences in the abundance of mites on Blackcaps, house sparrows, sardinian and willow warblers. This study was carried out just before the breeding season, thus the juveniles may have been “mite-contaminated” by adults during the winter. Also, blackcaps could potentially be carrying different mite species, collected enroute during their migration. With more observational data over different time of day and seasons, investigations could be carried out to describe mite movements depending on varying environmental factors.
Feather mites are a taxonomically diverse group of arthropods that live permanently on birds and represent the most common type of bird-mite associations (Proctor and Owens, 2000). Feather mites not only possess several morphological adaptations for living on birds but exhibit a range of specific behavioural responses to the constantly changing environmental conditions on wing feathers (Jovani and Serrano, 2004; Proctor, 2003). While mites have the ability to potentially inhabit all feathers of the wing - especially in the structurally similar wing feathers of passerines - they are known to have very clear microhabitat preferences (Bridge, 2003; Proctor, 2003; Thompson et al., 1997). Feather mites are sensitive to a number of environmental variables and their distribution on wings vary with conditions (e.g. moulting feathers, temperatures, direct sunlight), often resulting in significant seasonal, daily, or short-term shifts (Proctor, 2003).
The exact causes of differences in mites abundance and distribution is currently debatable (Proctor and Owens, 2000) as there is little evidence to prove high parasite load as a cause or consequence of poor host physiological condition in free-living birds (Thompson et al., 1997). Many studies have linked mite abundance to factors such as sex (e.g. Blanco et al., 1999), age (Hamstra and Badyaev, 2008), body condition (uropygial gland size, weight, fat levels, muscle scores, etc.) (Galvan and Sanz, 2006; Hartup et al., 2004; Harper, 1998), migratory habits (e.g. Moller et al., 2004), or habitat (e.g. Galvan and Sanz, 2006) but no consensus has been reached on underlying mechanisms. Part of the problem arises from the general lack of knowledge about feather mite biology and basic physiological and ecological effects on birds (Proctor and Owens, 2000).
The current lack of consensus for both parasitic and non-parasitic roles of feather mites calls for more studies describing bird-mite relationships and elucidating the functional significance of feather mites (Bridge, 2003). Previous studies (e.g. Gaud and Atyeo, 1996) found that feather mites can be associated to true parasites, thus correlational studies of birds and feather mites needs to be interpreted with caution.
1 Aims and Objectives
This study aimed to investigate if mite loads differ across species and to test if factors, such as age, sex and body condition could explain trends in mite abundances. It is hypothesized that feather mite density will be positively associated with poor body condition and that mite loads are affected by sex and age in a species. In addition, this paper investigated the preferences for certain microhabitats on a wing and by roosting several birds overnight, this study examines how mite distributions change over time as a result of changing environmental conditions/bird activity.
2 Methods and Analysis
Study area- Data was collected from 31st of March until 4th of April 2011 at Urra Field station near Sorbas, in Almeria province, south-east Spain. Annual rainfall is low during all seasons (ca 300 mm), mean temperatures of 17 ℃ but regularly rise to over 40 ℃. The area has been classified as semi-arid thermomediterranean (Rivas-Martinez et al., 2001) and is considered as the driest place in Europe.
Sampling- Birds were captured using mist nets around the Urra Field centre by authorised bird ringers. Nets were set when birds were most active (6 am~12 pm and 5~10 pm). In addition to the standard data collected by the ringers (species, sex, age, weight, tail and wing measurements), the following information was recorded: body condition, weight, fat and pectoral muscle levels were used as a proxy for body condition for each bird. Pectoral muscles and fat levels were scored for each bird, following BTO data collection and biometrics standards (2001) (See appendix 1 and 2, respectively).
|
Appendix 1 Scoring of fat levels
|

|
Appendix 2 Feather mite distributions across different areas of the wing for passerine species
|
Mite distribution on the wing-Feather mites on primary and secondary feathers was counted using handheld magnifying glasses. Mites found on the right wing were used in the final analysis because an initial comparison of feather mite abundances on the left and right wings showed that abundances were highly correlated (Pearson correlation R=0.975, df=71, p<0.001). The spatial distribution of mites on the right wing was recorded by dividing the wing into several categories: 1=primaries 1-3 (P1-3), 2=primaries 4-6 (P4-6), primaries 7-10 (P7-10), 4=secondaries, and 5=random distribution. Scores were assigned to each wing, depending on where more than 50% of the mites were found.
Diurnal changes in mite abundances-A number of birds were roosted overnight and mite numbers were counted both in the evening and on the following morning to determine changes over time in response to changing environments. Percentage change in mites overnight was calculated for each bird by using the following formulae:
M=[(x-y)/y]*100%
Where m is the percentage change
X is the evening mite abundance
Y is the morning mite abundance
Mite abundance data was non-normal so non-parametric statistics (Kruskal-Wallis test) were used to test for differences in mite abundance across species, sex, age and to test for differences in mite distribution on wings across species. Spearman’s correlations were used to test for relationships between mite abundances and body condition (weight, muscle and fat levels). Fisher’s exact test was used to test for changes in mite abundances on birds kept overnight to test observed vs. expected results (expected results being random distribution on the wing and no movement overnight respectively). In the case of the diurnal activity study, a 10% sampling error was assumed, as counting mites in low light conditions increases the risk of observer error. This estimate can be considered as highly conservative, as counting errors were generally very low (pers. obs.) even when mite abundances were high. Statistical analysis was restricted to the species with N≥10.
3 Results
A total of 132 birds (including 13 recaptures) from 20 different species were caught and examined for feather mites. Without considering recaptured birds, blackcap, Sardinian warbler, chiffchaff, willow warbler and house sparrow were the most commonly captured birds (Table 1), contributing to over 77% of the birds caught. Table 1 shows mite prevalence, average mite numbers and the number of birds caught per species. Most species had N<10 and were not used for analysis and although chiffchaff, willow warbler, and house sparrow were included in the analysis, the data should be treated with a degree of caution since samples size is small.
|
Table 1 Prevalence and average abundance (per wing) of mites in different bird species captured at Urra field centre, south-east Spain from 31st March-4th April 2011
|
.png)
The four most common species and showed a significant difference in mite abundances (Kruskal Wallis c2=31.193, df=3, p<0.001) as shown in Figure 1. Blackcaps had the highest average mite abundances, followed by house sparrows, Sardinian warblers and willow warblers. Both Sardinian and willow warblers were characterized by relatively low mite abundances but had a few outliers.
|
Figure 1 Feather mite abundances for the four most common species
|
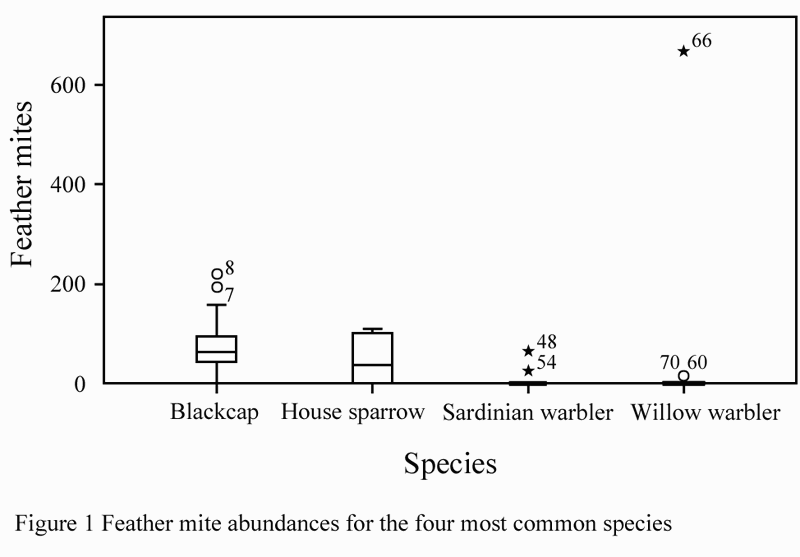
There are no significant differences in mite abundance between males and females in blackcaps (Kruskal Wallis c2=0.38, df=1, p=0.538); house sparrow (Kruskal Wallis c2=0.046, df=1, p=0.829); Sardinian warbler (Kruskal Wallis c2= 0.011, df=1, p=0.916) and willow warbler (Kruskal Wallis c2= 0.327, df=1, p=0.568).
No significant differences were found in mite abundance between juvenile and adults blackcap (Kruskal Wallis c2=0.012, df=1, p=0.912) and Sardinian warbler (Kruskal Wallis c2=0.359, df=1, p=0.549). It was not possible to analyse if age has an effect on mite abundances of willow warblers and house sparrows, as it was impossible to determine ages based on moulting patterns.
A summary of Spearman’s rank correlations of feather mites to body condition measures is given in Table 2. There were no significant correlations between feather mites with body conditions (fat, pectoral muscles and weight) at the p=0.05 significance level, except in Sardinian warbler where mites were positively correlated to fat levels (Spearman’s ρ=0.779, df=14, p<0.001); and in blackcap mites were marginally positively correlated to weight (Spearman’s ρ=0.296, df=35, p=0.085).
|
Table 2 Correlations of mite abundance to different measures of body condition
|
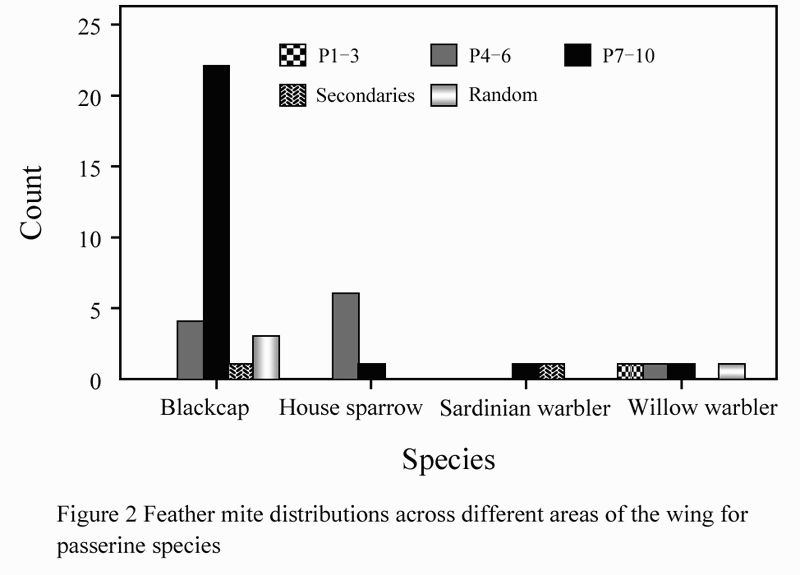
The occurrence of feather mites on wing regions (P1-3, P4-6, P7-10, secondaries and random distribution) varied significantly between the 4 species (Kruskal Wallis c2=12.388, df=3, p=0.006). As shown in Figure 2, mites on blackcap were mainly found on P7-10 (Fishers Exact test value =20.215, p<0.001) and mites on house sparrows were concentrated on P4-6 (Fishers Exact test value = 6.223, p<0.045).
|
Figure 2 Feather mite distributions across different areas of the wing for passerine species
|
There was significant reductions in the numbers of mites on the wings for birds roosted overnight (Fisher’s exact test value=6.223, p=0.045), indicating that there were significant daily changes in abundance on the wings and that these were greater than could be expected by sampling error alone.
4 Discussions
As these results clearly show, mite abundance varies significantly between different bird species, with some species (e.g. cirl buntings, chiffchaffs, greenfinches) having very high numbers of mites per wing. Average mite abundances can be misleading as some of the averages were influenced by outliers (e.g. chiffchaff). Considering bird species with both high prevalence of mite infestations and high average mite abundances, it becomes apparent that many of the more heavily and commonly infested bird species tend to be gregarious species, such as cirl bunting, greenfinches, and serin. These species often form large flocks, feeding and roosting together (Svensson et al., 1999), providing ideal opportunities for mites infestations (Proctor, 2003; Proctor and Owens, 2000). Species characterised by both low prevalence and low mite numbers tend to be solitary species (e.g. robins, Sardininan warblers, Bonelli’s warblers, woodchat shrikes), often occurring in pairs and highly territorial, with little or no contact to conspecifics. Although few birds were captured to carry out meaningful analysis, these trends concur with previous findings by Poulin (1991), who found a positive relationship between mite abundances and host gregariousness.
Blackcaps, which were among the most heavily infested species (100% prevalence, average 72.5 mites per wing), appear to be an exception to the trendas they tend to be relatively solitary, feeding and breeding in pairs and defending their territories against conspecifics. As suggested by Fowler and Hodson (1991), migratory birds such as blackcaps could potentially be carrying different mite species, collected enroute during migration.
4.1 Mite abundance in relation to sex and age
Various authors have suggested that mite abundances can be related to sex (Hamstra and Badyaev, 2008; Blanco and Frias, 2001; Blanco et al., 1999), mostly as a result of variations in hormone levels between different sexes (Blanco et al., 1999). Concurring with the findings from Galvan and Sanz (2006), Blanco et al (1997) and many others, the results of this study failed to find such associations for any of the species, indicating that the proposed variations in hormonal levels may not be strong enough to explain differences in mite abundances. While this may in part be due to the low number of captured birds, it is more likely that both sexes do indeed harbour similar abundances of mites.
Similarly, the results suggest no differences between mite abundances between juvenile and adult birds. Several studies found age to be important factor in determining mite abundances on birds, with mite loads usually increasing with increasing age (Hamstra and Badyaev, 2008; Blanco and Frias, 2001), attributed to the fact that mites are generally transferred from the female to the fledglings in the nest and then increase with reproduction and sociality over time (Galvan and Sanz, 2006;Figuerola, 2000; Poulin, 1991). As this study was carried out just prior to the breeding season, any effect of age may have been masked by the fact that juveniles have been exposed to infested adults throughout winter (high winter sociality) as suggested by McClure (1989).
4.2 Mite infestation and host body condition
The various scores of body condition used by this study were poor predictors of mite abundances. For the four species that were studied in more detail, only Sardinian warblers seemed to be characterized by a significant relationship between fat scores and mite loads (p<0.001), while the relationship between weight and feather mite load in blackcaps was marginally non-significant (p=0.085). The fact that both of these relationships were positive (as body condition improved, mite abundances increased) indicates that mites may not be detrimental to their hosts, contrary to what has been suggested by many authors (e.g. Figuerola et al., 2002; Figuerola, 2000; Harper, 1999; Thompson, 1997) . Whether good body condition of the host provides mites with ideal body conditions (thus explaining the high abundances) or if this trend was cause by underlying covariates was beyond the scope of this paper and would require manipulative/experimental rather than observational studies. Future research into this area could provide vital insights into the mechanisms underlying mite-host relationships and the effects of mites on their avian hosts.
4.3 Mite distribution on wings and movements
The observed significant mite distributions on species could be explained by the microhabitat preferences that may be different for the mite species, (Galvan et al., 2007) some which are host specific. It has already been discovered that mites might migrate on the feathers depending on environmental factors such as temperature (e.g. Proctor, 2003). Such migrations are likely to have caused the observed trends in the birds that were roosted overnight. With more observational data over different time of day and seasons, investigations could be carried out to describe mite movements depending on varying environmental factors (e.g. temperature, sunlight, rain).
Reference
Behnke J., McGregor P., Cameron J., Hartley I., Sheperd M., Gilbert F., Barnard C., Hurst J., Gray S., and Wiles R., 1999, Semi-quantitative assessment of wing feather mite (Acarina) infestations on passerine birds from Portugal. Evaluation of the criteria for accurate quantification of mite burdens, Journal of Zoology, 248(3): 337-347
http://dx.doi.org/10.1111/j.1469-7998.1999.tb01033.x
Blanco G., and Frias O., 2001, Symbiotic feather mites synchronize dispersal and population growth with host sociality and migratory disposition, Ecography, 24(2): 113-120
http://dx.doi.org/10.1034/j.1600-0587.2001.240201.x
Blanco G., Seoane J., and de la Puente J., 1999, Showiness, non-parasitic symbionts, and nutritional condition in a passerine bird, Ann. Zool. Fennici, 36: 83-91
Bridge E.S., 2003, Densities and distributions of commensal feather mites (Zachvatninia caspica) among the primaries of caspian terns, International Journal of Acarology, 29: 389-398
http://dx.doi.org/10.1080/01647950308684356
British Trust for Ornithology (BTO), 2001, Data collection and biometrics, British Trust for Ornithology,UK
Figuerola J., 2000, Ecological correlates of feather mite prevalence in passerines, Journal of Avian Biology, 31(4): 489-494
http://dx.doi.org/10.1034/j.1600-048X.2000.310408.x
Figuerola J., Domenèch J., and Senar J.C., 2002, Plumage colour is related to ectosymbiont load during moult in the serin, Serinus serinus: an experimental study, Animal Behaviour, 65(3): 551-557
http://dx.doi.org/10.1006/anbe.2003.2072
Fowler J.A., and Hodson D.P., 1991, Feather lice (Mallophaga) of the dipper Cinclus cinclus in central Wales, Ringing and Migration, 12(1): 43-45
http://dx.doi.org/10.1080/03078698.1991.9673983
Galvan I., and Sanz J.J., 2006, Feather mite abundance increases with uropygial gland size and plumage yellowness in great tits Parus major, Ibis, 148(4): 687-697
http://dx.doi.org/10.1111/j.1474-919X.2006.00576.x
Galvan I., Barba E., Piculo R., Cantó J.L., Cortés V., Monrós J.S., Atiénsar F., and Proctor H., 2007, Feather mites and birds: an interaction mediated by uropygial gland size? Journal of Evolutionary Biology, 21(1): 133-144
PMid:18028353
Gaud J., and Atyeo W.T., 1996, Feather mites of the world (Acarina, Astigmata): the supraspecific taxa, Ann. Sci. Zool., 277: 1-193
Hamstra T.L., and Badyaev A.V., 2008, Comprehensive investigation of ectoparasite community and abundance across life history stages of avian host, Journal of Zoology, 278(2): 91-99
http://dx.doi.org/10.1111/j.1469-7998.2008.00547.x
Harper D.G.C., 1999, Feather mites, pectoral muscle condition, wing length and plumage coloration of passerines, Animal Behaviour, 58(3): 553-562
http://dx.doi.org/10.1006/anbe.1999.1154 PMid:10479371
Hartup B.K., Stott-Messick B., Guzy M., Ley D.H., 2004, Health survey of house finches (Carpodacus mexicanus) from Wisconsin, Avian Diseases, 48(1): 84-90
http://dx.doi.org/10.1637/7067 PMid:15077801
Jovani R., and Serrano D., 2004, Fine-tuned distribution of feather mites (Astigmata) on the wing of birds: the case of blackcaps Sylvia atricapilla, Journal of Avian Biology, 35(1): 16-20
http://dx.doi.org/10.1111/j.0908-8857.2004.03213.x
McClure H.E., 1989, Occurrence of feather mites (Proctophyllodidae) among birds of Ventura county lowlands, California, Journal of Field Ornithology, 60(4): 431-450
Moller A.P., de Lope F., and Saino N., 2004, Parasitism, immunity, and arrival date in a migratory bird, the barn swallow, Ecology, 85(1): 206-219
http://dx.doi.org/10.1890/02-0451
Poulin R., 1991, Group-living and infestation by ectoparasites in passerines, The Condor, 93: 418-23
http://dx.doi.org/10.2307/1368958
Proctor H.C., 2003, Feather mites (Acari: Astigmata): ecology, behaviour, and evolution, Annual Review of Entomology, 48: 185-209
http://dx.doi.org/10.1146/annurev.ento.48.091801.112725 PMid:12208818
Proctor H., and Owens I., 2000, Mites and birds: diversity, parasitism and coevolution, Tree, 15(9): 358-364
http://dx.doi.org/10.1016/S0169-5347(00)01924-8
Rivas-Martinez S., Penas A., and Diaz T.E., 2001, Biogeographic map of Europe, Mapping Service, University of León, León, Spain
Svensson L., Grant P.J., Mullarney K., and Zetterström D., 1999, Collins bird guide: the most complete guide to the birds of Britain and Europe, Harper Collins Publishers, London, UK
Thompson C.W., Hillgarth N., Leu M., and McClure H.E., 1997, High parasite load in house finches (Carpodacus mexicanus) is correlated with reduced expression of a sexually selected trait, The American Naturalist, 149(2): 270-294
http://dx.doi.org/10.1086/285990



. PDF(177KB)
. FPDF
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Tawanda Tarakini
. Mikis Bastian
Related articles
. Feather mites
. Pectoral muscle
. Body weight
. Fat condition
. Mite hosts
Tools
. Email to a friend
. Post a comment