The Effects of Diverse Habitats in Urra on Lizard Abundance 
 Author
Author  Correspondence author
Correspondence author
International Journal of Molecular Evolution and Biodiversity, 2011, Vol. 1, No. 1 doi: 10.5376/ijmeb.2011.01.0001
Received: 14 Nov., 2011 Accepted: 28 Nov., 2011 Published: 08 Dec., 2011
Colomba and Liang, 2011, The effects of diverse habitats in Urra on lizard abundance, International Journal of Molecular Evolution and Biodiversity, Vol.1, No.1 (doi: 10.5376/ijmeb.2011.01.0001)
Lizards prefer certain habitat in terms of their morphological traits. Lizard abundance is an important indicator of their habitat suitability. Besides, foraging and basking are important behaviours for lizards, which largely depend on the habitat types. Psammodromus (Psammodromus algirus) and Spanish psammodromus (Psammodromus hispanicus) mainly inhabit in the Iberian Peninsula. The gypsum deposits distribute throughout this region and small proportion of the area are sandy river valley. This research is implemented on the gypsum deposits and sandy river valley with different vegetation distribution. We would like to figure out the effect of diverse habitats on lizard abundance. It may be influential in population ecology as well as biodiversity of lizards and their habitat.
Lizards in particular have specific morphological features, which are adaptive to their certain preferred habitat (Martín and Salvador, 1992). Hence, species abundance is an indicator of habitat suitability (Diaz and Carrascal, 1991). If the variables are known that determine distinct habitats, then testing which of these factors are most important to lizard species abundance and population densities would yield valuable results for population ecology and biodiversity conservation (Case, 1975).
The Spiny-footed lizard (Acanthodactylus erythrurus), large Psammodromus (Psammodromus algirus) and Spanish Psammodromus (Psammodromus hispanicus) are all medium-sized ground dwelling lacertid lizards (Arnold, 1987; Díaz and Carrascal, 1991). They are found predominately in the Iberian Peninsula (Martín and López, 2003) at elevations up to 1,000m, but are usually more abundant lower (Arnold and Ovenden, 2002). A. erythrurus prefers a wide variety of habitats, such as open, sandy areas with sparse vegetation distribution (Arnold, 1987). They feed on ants (Perez-Mellado, 1992; Pérez-Quintero, 1996) and some active prey (Belliure et al., 1996). P. algirus can be found in open or degraded woodland, and eucalyptus forest (Arnold and Ovenden, 2002), while P. hispanicus is found in the sandy soils with low dense, bushy plants (Arnold and Ovenden, 2002). Small arthropods are Spanish Psammodromus main food source (Arnold and Ovenden, 2002).
Foraging and basking are important behaviours for lizards. The Psammodromus species capture food items mainly in the leaf litter underneath shrubs layer; sometimes a short foraging expedition happens due to the active prey (Diaz and Carrascal, 1993). However, A. erythrurus is definitely a less energetic predator (Belliure et al., 1996). It spends more time sitting and waiting. Moreover, lizards may also prefer rocky habitats, where they can avoid predators in crevices and bask simultaneously using these rocks as perches. Lizards bask to absorb the sun until they reach the optimum temperature in order to carry out their daily activities (Martín and Salvador, 1995). If lizards occupy more open habitats, they will increase their basking frequency (Schwartz and Thomas, 1975). Also, rocky slopes maximize heating rate, which in turn minimizes overall basking time. Therefore, lizards have more time to do other activities such as mate or forage (Martín and Salvador, 1995). Moreover, lizards need refuge to run into to avoid predators and the escape distances are relative to the vegetation cover because it makes up for the bare ground.
In our study, we tested the effects two diverse habitats had on P. algirus, P. hispanicus, A. erythrurus. We hypothesize that lizards will be more active at times when the sun is directly hitting the surface, and lizards will prefer open areas within a close distance to low shrub cover, to maximize their opportunities to bask, forage and avoid predators.
1 Results
1.1 Escape distance
The correlation between escape distance and temperature was shown to be non significant (ρ=0.095, n=46, P=0.529), and escape distance and wind speed was also not significant (ρ=0.081, n=46, P=0.594). The Kruskal-Wallis test also showed that escape distance was not significant (χ2=0.433, df=3, P=0.933; Figure 1). The Mann-Whitney Test showed the relative escape distance is almost the same (U=141.5, n=42, P=0.283).
|
Figure 1 The escape distance of A. erythrurus and Psammodromus sp.
|

1.2 Lizard abundance
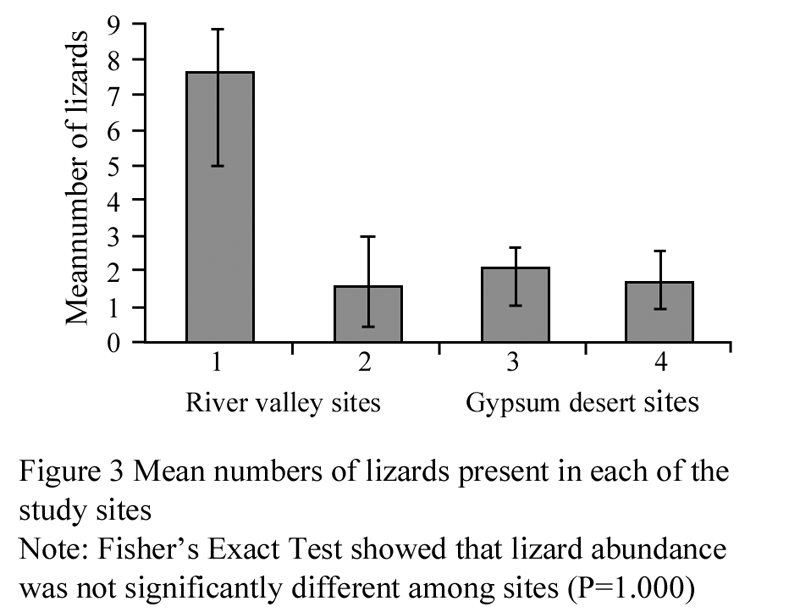
Using the Fisher’s Exact Test, aspect was significantly related the number of lizards that were found on the side that the sun was shining (P<0.001). Similarly, the Fisher’s Exact Test gave a significant result for lizard abundance in each site, separated by time (P<0.001; Figure 2). Conversely, the Fisher’s Exact Test gave non-significant results for the number of lizard species (A. erythrurus and Psammodromus sp.) found in each site (P=1.000). According to the Kruskal-Wallis Test, lizard abundance was not significantly different at each of the different sites (χ2= 5.115, df = 3, P= 0.164; Figure 3).
|
Figure 2 The number of A. erythrurus and Psammodromus sp.
|
.png)
|
Figure 3 Mean numbers of lizards present in each of the study sites
|
1.3 Habitat variables
The percent vegetation cover was not significantly different per site (χ2=2.412, df=3, P=0.491; Figure 4). The Generalized Linear Model for the effects the interactions of habitat, wind speed and temperature had on escape distance showed all non significant results. There was no minimum adequate model because even when the two way interaction terms were omitted individually (wind speed × air temperature; wind speed × habitat; air temperature × habitat), to reveal just the main effects, the model was still insignificant (habitat: Wald χ2=0.049, df=1, P=0.849; air temperature: Wald χ2=0.437, df=1, P=0.509; wind speed: Wald χ2=0.582, df=1, P=0.445).
|
Figure 4 The mean percent of vegetation cover per site
|
.png)
2 Discussions
2.1 Escape distance
Since lizard abundance and vegetation cover did not differ significantly between sites, this indicates that lizard abundance is not related to vegetation cover. This is surprising because Stamps (1983) showed that vegetation structure played a critical role in reptile habitat selection for predator escape. Additionally, this vegetation cover has also been used for foraging (Karasov and Anderson, 1984), and mating (Diaz and Carrascal, 1991). Algyroides marchi is a lizard endemic to the mountainous region of south-east Iberian Peninsula. The more stoney and vegetated the environment, then the less abundant they are (Rubio and Carrascal, 1994). This could relate to A. erythrurus and Psammodromus sp., which could explain why lizard abundance was not correlated with vegetation cover (Figure 3; Figure 4).
The habitats did not differ in percent vegetation cover (Figure 1). This is supported by previous research; both P. algirus (Diaz and Carrascal, 1991) and A. erythrurus population densities are correlated with leaf litter and low dense shrub cover (Martín and López, 2003), which was available in similar amounts on all of our sites. Hence, this is also why the Mann-Whitney Test showed that lizard species did not differ in their relative escape distances.
2.2 Lizard abundance
One would expect morning counts of lizards to be greater than in the afternoon. In the morning, A. erythrurus were more active in the river valley, while in the afternoon, were more active in gypsum desert (Figure 2). Busack (1976) showed that while A. erythrurus activity peaks in the morning, adults stay in cool microhabitats but within their thermal limits, which allow them to remain active throughout the day. A. erythrurus was the only species found on April 3, a cloudy day when the air temperature was lower and the relative humidity was higher, which support Busack (1976)’s results. In contrast, P. hispanicus cannot raise their body temperatures on cloudy days via basking, and hence are unable to forage for long periods of time (Patterson and Davies, 1984), which may explain why they were not seen on April 3. Additionally, A. erythrurus could be found in the river valley in the morning. However, it should be noted that there was a high chance of duplicate sightings of the same individual as we were unable to mark the species we documented.
Given the desert’s natural sloping landscape, this could have been a reason why more lizards were found there (Figure 2). Likewise, the aspect was also significant, as nearly all of the lizards (n=44) were sighted where the sun was hitting the ground, and only (n=3) were not. Bohorquez-Alonso et al (2011) discovered Gallotia galloti, another Lacertid, were mostly found oriented parallel or perpendicular to the sun. Also, Arribas (2010) studied activity and microhabitat selection between two Pyrenean rock lizards, both Lacertids found in Spain. He discovered that Iberolaceria aurelioi were found on steeper areas than I. bonnali and I. aranica. Further studies should be conducted analyzing slope together with aspect to determine if the sun is hitting the slopes in the gypsum desert more directly in the afternoon than the evening, which would explain our result.
The Fisher’s Exact Test showed that the different species of lizards had no preference for a particular habitat. One possible explanation could be because they have evolved adaptations for both environments. For example, another lizard found in our study area, the ocellated lizard (Lacerta lepida) is found in warm, arid climates, and also in wet, cold regions of Spain (Hódar et al., 1996). These traits could suggest a similar adaptation for A. erythrurus, P. hispanicus and P. algirus. However, if we were to quantify the other habitat variables (presence of rocks, soil composition, and prey abundance) and run further analyses then possibly our results would yield different outcomes, as our two habitats may show to be more diverse. Likewise, Michael et al (2010) discovered that rocky environments are related to reptile abundance.
The Generalized Linear Model showed that the interaction between habitat, wind speed and air temperature had no effect one escape distance. In future experiments, it would be wise to take calculations of temperature and wind speed even at sites where no lizards are found. Then a Generalized Linear Model can be constructed on the effect of these interactions terms, together with habitat and escape distance on lizard abundances.
To conclude, lizards did not have a preference for one particular habitat since there was no difference between the escape distance, vegetation cover and, therefore, no difference between lizard abundances between each of the sites. This implies that these three species are occupying the same space, and feeding on the same arthropod populations and foraging in the same area. Thus, species competition to determine if these species have (or potentially will) evolve certain adaptations to the environment needs further studies.
3 Methods
3.1 Study area
Observations were conducted during spring (April 1-4, 2011) in the sparsely vegetated habitats of the sloppy desert in Sorbas Karst Gypsum Natural Monument, and in a sandy (Hughes, 2011, unpublished data) dry river valley near Sorbas, Almeria Province, South-east Spain. Dominant vegetation in the region is Stipa Tenacissima, Anthyllis cytisoides, Retama Sphaerocarpa, Garigue shrubs, and tall maquis-like vegetation in the river valley (Hughes, 2011, unpublished data).
3.2 Field recordings
Between the hours of 0930-1330 and 1430-1730 hours (G.M.T.+1), we haphazardly walked at a constant pace of about 15m per minute in a zigzag pattern (Martín and López, 2003) for 1.5 hours in each of two study sites (20m apart) in both the gypsum desert and the dry river valley. Approximately every four minutes on the first day, a 1m2 quadrat sample was taken of percent vegetation cover. Twenty quadrats were taken in total for each site. At every lizard sighting, the date, time, identification, size, and color of species, adult/juvenile, microhabitat with presence of prey, air temperature/humidity of sunny/shaded areas, escape distance to refuge, wind speed (Schoener, 1970; Pianka and Parker, 1972), and aspect were recorded. Digital photographs aided in further identification of lizard and vegetation species. A compass was used to record the aspect (Fisher et al., 2003), and a thermo-hydro-anemometer was used to record the temperature, humidity and wind speed. A transect line was used to measure the escape distance from the start point (where we first spotted the lizard) to the refuge.
4 Data analysis
All of the raw data showed non-normal distribution, which when tested with a Shapiro-Wilks normality test for both logged and square root transformed data still showed non-normal distribution. Therefore, non-parametric tests were rendered. The differences on habitats were analyzed via Fisher’s Exact Test. A Pearson’s correlation between escape distance and temperature, and also escape distance and wind speed were analyzed. Mann-Whitney Test was used to test for a significant difference between each species in their relative escape distances. Kruskal-Wallis tests were used to analyze the difference of lizard numbers for each site; the vegetation cover for each site; and escape distance for each site. Fisher’s Exact Test was performed to dictate the abundance of each species in each habitat, and also to test for significance of lizard abundances in each habitat as a function of time. A Generalized Linear Model was constructed to test for significance of the effect of the interactions between wind speed × air temperature × habitat (and all of the 2 way interactions) on escape distance. Data analyses were performed using SPSS 17 and Microsoft Excel 2007.
References
Andersson M., Krockenberger A., and Schwarzkopf L., 2010, Experimental manipulation reveals the importance of refuge habitat temperature selected by lizards, Austral Ecology, 35: 294-299
http://dx.doi.org/10.1111/j.1442-9993.2009.02035.x
Arnold E.N., 1987, Resource partitioning among lacertid lizards in Southern Europe, J. Zool. Lond, 1: 739-782
Arnold N., and Ovenden D., 2002, A field guide to the reptiles and amphibians of Britain and Europe, Harper Collins Publishers, London, United Kingdom
Arribas O.J., 2010, Activity, microhabitat selection and thermal behavior of the Pyrenean Rock Lizards Iberolacerta aranica (ARRIBAS, 1993), I. aurelioi (ARRIBAS, 1994) and I. bonnali (LANTZ, 1927), Herpetozoa, 23: 3-23
Bauwens D., 1981, Survivorship during hibernation in the European common lizard, Lacerta vivipara, Copeia, 3: 741-744
http://dx.doi.org/10.2307/1444592
Belliure J., Carrascal L.M., and Diaz J.A., 1996, Covariation of thermal biology and foraging mode in two Mediterranean lacertid lizards, Ecology, 77(4): 1163-1173
http://dx.doi.org/10.2307/2265585
Bohorquez-Alonso M.L., Font E., and Molina-Borja M., 2011, Activity and body orientation of Gallotia galloti in different habitats and daily times, Amphibia-Reptilia, 32(1): 93-103
http://dx.doi.org/10.1163/017353710X542994
Busack S.D., 1976, Activity cycles and body temperatures of Acanthodactylus erythrurus, Copeia, 4: 826-830
http://dx.doi.org/10.2307/1443478
Case T.J., 1975, Species numbers, density compensation, and colonizing ability of lizards on islands in the Gulf of California, Ecology, 56(1): 3-18
http://dx.doi.org/10.2307/1935296
Diaz J.A., and Carrascal L.M., 1991, Regional distribution of a Mediterranean lizard: influence of habitat cues and prey abundance, J. of Biogeography, 18(3): 291-297
http://dx.doi.org/10.2307/2845399
Diaz J.A., and Carrascal L.M., 1993, Variation in the effect of profitability on prey size selection by the lacertid lizard Psammodromus algirus, Oecologia, 94(1): 23-29
http://dx.doi.org/10.1007/BF00317296
Fisher R.N., Suarez A.V., and Case T.J., 2002, Spatial patterns in the abundance of the Coastal Horned Lizard, Conservation Biology, 16(1): 205-215
http://dx.doi.org/10.1046/j.1523-1739.2002.00326.x
Hódar J.A., Campos F., and Rosales B.A., 1996, Trophic ecology of the ocellated Lizard Lacerta lepida in an arid zone of southern Spain: relationships with availability and daily activity of prey, J. of Arid Environments, 33: 95-107
http://dx.doi.org/10.1006/jare.1996.0049
Karasov W.H., and Anderson R.A., 1984, Interhabitat differences in energy acquisition and expenditure in a lizard, Ecology, 65(1): 235-247
http://dx.doi.org/10.2307/1939476
Martín J., and López P., 2003, Changes in the escape responses of the lizard Acanthodactylus erythrurus under persistent predatory attacks, Copeia, 2003(2): 408-413
Martín J. and Salvador A., 1992, Tail loss consequences on habitat use by the Iberian Rock Lizard, Lacerta monticola, Oikos, 65(2): 328-333
http://dx.doi.org/10.2307/3545026
Martín J., and Salvador A., 1995, Microhabitat selection by the Iberian Rock Lizard Lacerta monticola: effects on density and spatial distribution of individuals, Biological Conservation, 79(2-3): 303-307
Menezes V.A., Sluy M., Fontes F.A., and Rocha C.F.D., 2011, Living in a caatinga-rocky field transitional habitat: ecological aspects of the whiptail lizard Cnemidophorus ocellifer (Teiidae) in northeastern Brazil, Zoologia (Curitiba, Impr.), 28(1): 8-16
Michael D.R., Cunningham R.B., and Lindenmayer D.B., 2010, Microhabitat relationships among five lizard species associated with granite outcrops in fragmented agricultural landscapes of south-eastern Australia, Austral Ecology, 35(2): 214-225
http://dx.doi.org/10.1111/j.1442-9993.2009.02029.x
Patterson J.W., and Davies P.M.C., 1984, The influence of temperature, sexual condition, and season on the metabolic rate of the lizard Psammodromus hispanicus, Journal of Comparative Physiolofy B, 154(3): 311-316
http://dx.doi.org/10.1007/BF02464412
Pérez-Quintero J.C., 1996, Reproductive characteristics of three Mediterranean lizards: Psammodromus algirus (L), Psammodromus hispanicus Fitzinger and Acanthodactylus erythrurus (Schinz), Amphibia-Reptilia, 17(3): 197-208
http://dx.doi.org/10.1163/156853896X00388
Pianka E.R., and Parker W.S., 1972, Ecology of the iguanid lizard Callisaurus draconoide, Copeia, 1972(3): 493-508
http://dx.doi.org/10.2307/1442922
Rubio J., and Carrascal L.M., 1994, Habitat selection and conservation of an endemic Spanish lizard Algyroides marchi (Reptilia, Lacertidae), Biological Conservation, 70(3): 245-250
http://dx.doi.org/10.1016/0006-3207(94)90169-4
Santos X., and Poquet J.M., 2010, Ecological succession and habitat attributes affect the postfire response of a Mediterranean reptile community, European Journal Of Wildlife Research, 56(6): 895-905
http://dx.doi.org/10.1007/s10344-010-0387-8
Schoener T.W., 1970, Nonsynchronous spatial overlap of lizards in patchy habitats, Ecology, 51(3): 408-418
http://dx.doi.org/10.2307/1935376
Schwartz A., and Thomas R., eds., 1975, A checklist of West Indian amphibians and reptiles, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania, USA
Stamps J.A., 1983, Territoriality and the defense of predator-refuges in juvenile lizards, Animal Behaviour, 31(3): 857-870
http://dx.doi.org/10.1016/S0003-3472(83)80241-3



. PDF(173KB)
. FPDF
. HTML
. Online fPDF
Associated material
. Readers' comments
Other articles by authors
. Michelle Colomba
. Xingxing Liang
Related articles
. Psammodromus hispanicus
. Psammodromus algirus
. Lizards abundance
. Habitat
. Forage and bask
Tools
. Email to a friend
. Post a comment