Research Article
Genetic Diversity of Cultivated A. auricula by Comprehensive Analysis of rDNA-ITS and SRAP 
 Author
Author  Correspondence author
Correspondence author
International Journal of Molecular Evolution and Biodiversity, 2022, Vol. 12, No. 2 doi: 10.5376/ijmeb.2022.12.0002
Received: 14 Apr., 2022 Accepted: 27 Apr., 2022 Published: 08 May, 2022
Liu K.A., Ma H., Liu M., Zhang G.W., Yin S.L., and Li S.S., 2022, Genetic diversity of cultivated A. auriculala by comprehensive analysis of rDNA-ITS and SRAP, International Journal of Molecular Evolution and Biodiversity, 12(2): 1-10 (doi: 10.5376/ijmeb.2022.12.0002)
The genetic diversity of 15 A. auricula cultivars in North China was analyzed by strain affinity test, rDNA its and SRAP cluster analysis. The genetic relationship among the strains was clarified, which provided theoretical basis for the cultivation, genetic breeding and germplasm resources protection of A. auricula. The results showed that most of the antagonistic reactions of 15 A. auricula strains showed two kinds of incompatibility, i.e. non uplift type and ravine type, among which 5 pairs had no antagonistic reaction between A. auricula varieties, while ‘Jiyou No.1’ had obvious antagonistic reaction with other strains; rDNA its cluster analysis divided 15 A. auricula strains into three categories, the genetic distance between including ‘Heimuer-1’ and ‘Heimuer-2’, ‘Heiweibanjin’ and ‘Longjianghei+’, ‘Diaodai’ and ‘Heishan’ were 0, while that between ‘Jiyou No.1’ and other varieties was larger. The 9 SRAP primers amplified bands were rich in polymorphism, and the polymorphic fragments accounted for 79.8% of the total. The 15 cultivated species of A. auricula were divided into 3 categories by cluster analysis. The similarity coefficient among the strains of ‘Heiweibanjin’ and ‘Longjianghei+’ was 95%, which indicated that there was no genetic difference between them. The genetic relationship between ‘Jiyou No.1’ and other strains was far.
A. auricula (L.e. X Hook.) Underw. is also known as “Muer”, “Murong”, “Guangmuer” in Chinese. It is a large edible fungus, which is belongs to Auricularia genus in Auriculariaceae family, Auriculariales order, Hymenomycetes class, Basidiomycota subphylum, Eumycota phylum, Mycota kingdom (Shen, 2014). A. auricula is rich in a variety of amino acids and trace elements and elastic colloidal substances, containing functional nutrients such as polysaccharides, melanin, polyphenols, flavonoids, etc., can help digestion, prevent thrombosis, arteriosclerosis, iron deficiency anemia and coronary heart disease, etc., in addition to a certain anti-cancer effect. It is a nutritious edible fungus with pharmacological effects (Wang and Liu, 2014).
At present, there are many cultivated varieties of A. auricula, and the phenomenon of homonym and synonym is common. On the one hand, it may cause regional product simplification and increase the cultivation risk, and on the other hand, it is not conducive to the breeding and variety protection of A. auricula. DNA molecular markers are not affected by environment, tissue specificity, developmental stage and other factors, and can directly reflect genetic variation at DNA level (Du et al., 2013; Wang et al., 2019). Currently, molecular markers have been successfully applied in germplasm resources identification and genetic diversity evaluation of many plants and microorganisms (Yao et al., 2017; Wang et al., 2018). Sequence related amplified polymorphism (SRAP) is the specific amplification of ORFs (Open reading frames) region through forward and reverse primers. The forward and reverse primers can form primer pairs through free combination, which is universal, easy operation, high polymorphism and good repeatability. It has been applied to the evaluation of germplasm resources and genetic diversity analysis of various edible fungi such as A. auriculalar (Shi et al., 2019), Lentinula edodes (Liu et al., 2017; Wang et al., 2019), Grifola frondosa (Wang et al., 2019), Stropharia rugosoannulata (Zhu et al., 2018). The internal transcribed spacer (ITS) is the non-coding transcription spacer located between 18S and 5.8S for ITS1 and between 5.8S and 28S for ITS2 in the ribosomal rDNA. The internal transcribed spacer (ITS) is generally consistent within the species and significantly different between species. It can be used to analyze the phylogenetic relationship between bacteria with obvious differences between species within genus or within species (Jin et al., 2012; Zhang et al., 2013; Liu et al., 2015). Due to the different principles of each identification method, clustering analysis results are usually different. In order to ensure the accuracy of identification results, a variety of molecular marker methods are often used for comprehensive analysis (Shi et al., 2019).
In this study, the genetic diversity of 15 A. auricula cultivars was analyzed by rDNA-ITS and SRAP molecular markers combined with affinity test, and the genetic relationship between strains was clarified, providing theoretical basis for A. auricula cultivation, genetic breeding and germplasm resource protection.
1 Results and Analysis
1.1 Antagonistic reaction of different A. auricula cultivars
It is a traditional method to determine the affinities of A. auricula cultivated species by observing the antagonistic reaction of different varieties of mycelia contact to judge the genetic differences between strains. The results of affinity reaction of 15 A. auricula cultivars showed (Table 1) that 'Diaodai' and 'Heimuer 2', 'Heiweibanjin' and 'Longjianghei+', 'Longjianghei+' and 'Jipin-8', 'Heiweibanjin' and 'Jipin-8', 'Heimuer-2' and 'Heimuer-1' were somatic affinity, and other cultivars were not. In addition, 'Jiyou No.1' showed obvious antagonism with other strains (Figure 1). We preliminarily believed that 'Jiyou No.1' was different from the other 14 cultivated species of A. Auricula.
.png) Table 1 Antagonistic reaction of 15 strains of A. auricula Note: "+" represents incompatibility between strains; "-" represents affinity between strains, the strain number is the same as that in Table 3 |
.png) Figure 1 Antagonistic reaction between ‘Jiyou No.1’ and 14 other A. auricula strains |
1.2 Identification of A. auricula strains
DNA was extracted from 15 cultivated species of A. auricula for authenticity identification of the strain (Figure 2). ITS1 and ITS4 primers were used for PCR amplification of 15 A. auricula cultivars, and ITS sequences were sequenced to obtain ITS sequences. Phylogenetic tree was constructed by using proximity method (Figure 3). The 15 A. auricula cultivars were divided into three categories, among which the first category was' Jiyou No.1', 'Heimuer-1', 'Heimuer-2'. The second category is: 'Heishan', 'Diaodai', 'Heiweidanpian', 'Gaochanwang', 'Baolin No.2'; The third category is: 'Longjianghei+', 'Heiweijinban', 'Jupin-8', 'Yente No.5', 'Heiwei 15', 'Hei 29', and 'Heimuer 2'.
.png) Figure 2 Electrophoresis of 15 A. auricula strains genome DNA Note: M: Trans2K Plus II DNA Marker; 1~15: No. of A. auricula cultivar |
.png) Figure 3 Phylogenetic tree of 15 A. auricula strains based on ITS |
The genetic distance was calculated based on ITS sequences of 15 A. auricula cultivars (Table 2). The genetic distance between 'Heimuer-1' and 'Heimuer-2' strains in the first category was 0, indicating that there was no genetic difference between them. In the second group, the genetic distance between 'Diaodai' and 'Heishan' strains was 0, indicating that there was no genetic difference between them. In the third category, the genetic distance between 'Heiweibanjin', 'Longjianghei +' and 'Jipin-8' is 0, indicating that there is no genetic difference between them.
.png) Table 2 Genetic distance of 15 A. auricula strains based on ITS |
1.3 SRAP identification of A. auricula strain
Seventeen pairs of nucleotide primers were selected to amplify the sample mycelium DNA, and the strain numbers were the same as those (Table 3). Nine pairs of primers could amplify bands with clear bands and polymorphism (Figure 4; Figure 5; Figure 6; Figure 7; Figure 8; Figure 9; Figure 10; Figure 11; Figure 12), and the size of amplified fragments ranged from 100 to 2 000 bp. The number of DNA fragments amplified by each primer ranged from 4 to 14, with an average of 9.89. A total of 89 DNA fragments were amplified by the 9 primer pairs, and 71 different bands were amplified, and the polymorphic fragments accounted for 79.8% of the total, indicating that the amplified bands of the screened primers were rich in polymorphism and could be used for the analysis of genetic relationship.
.png) Table 3 Strains of A. auricular and their sources |
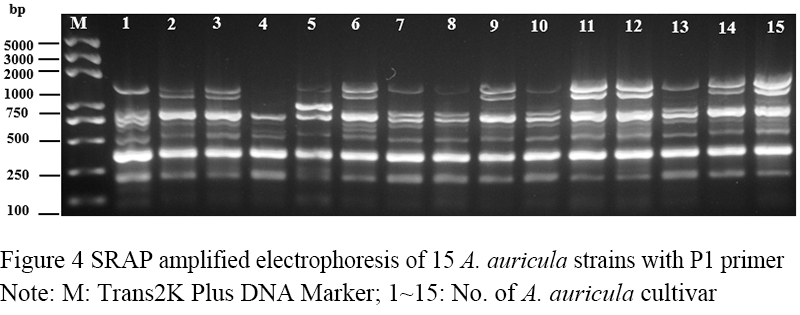 Figure 4 SRAP amplified electrophoresis of 15 A. auricula strains with P1 primer Note: M: Trans2K Plus DNA Marker; 1~15: No. of A. auricula cultivar |
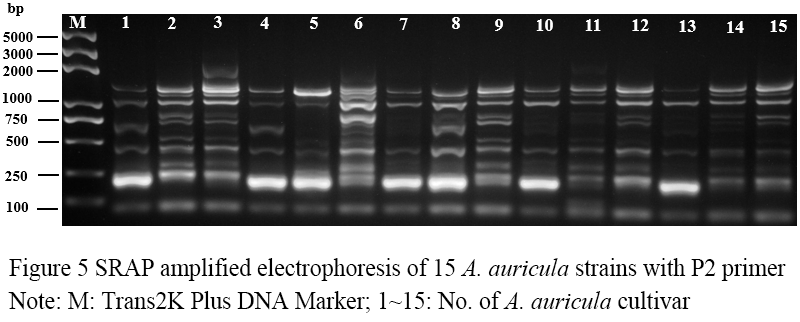 Figure 5 SRAP amplified electrophoresis of 15 A. auricula strains with P2 primer Note: M: Trans2K Plus DNA Marker; 1~15: No. of A. auricula cultivar |
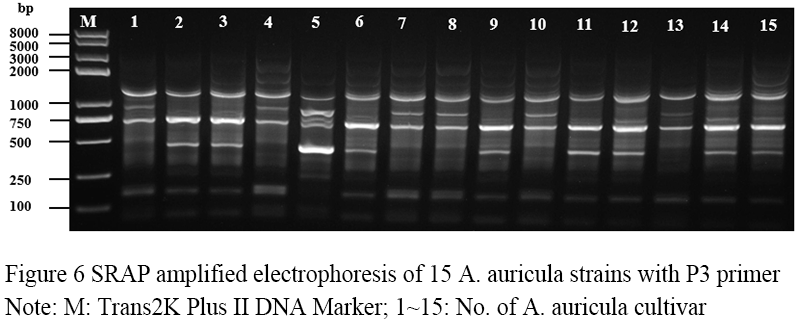 Figure 6 SRAP amplified electrophoresis of 15 A. auricula strains with P3 primer Note: M: Trans2K Plus II DNA Marker; 1~15: No. of A. auricula cultivar |
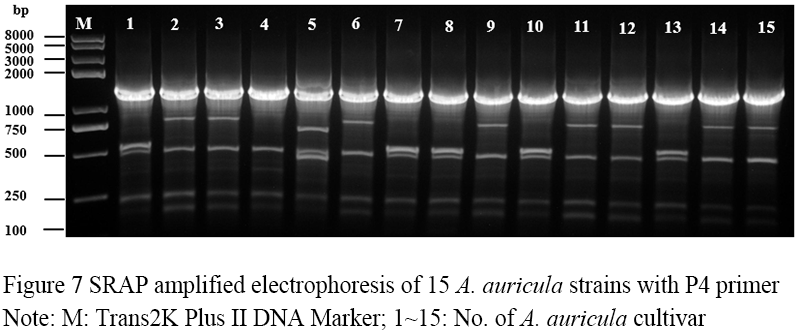 Figure 7 SRAP amplified electrophoresis of 15 A. auricula strains with P4 primer Note: M: Trans2K Plus II DNA Marker; 1~15: No. of A. auricula cultivar |
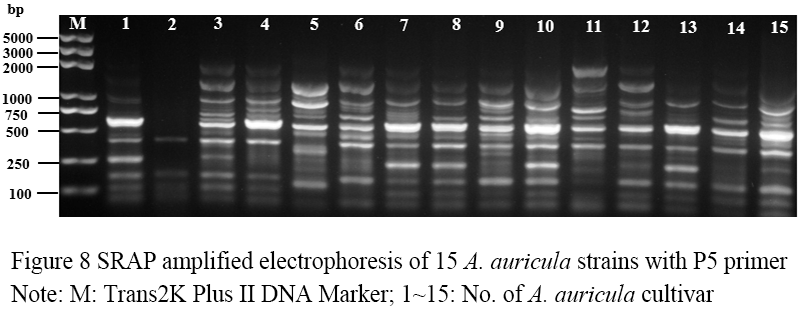 Figure 8 SRAP amplified electrophoresis of 15 A. auricula strains with P5 primer Note: M: Trans2K Plus DNA Marker; 1~15: No. of A. auricula cultivar |
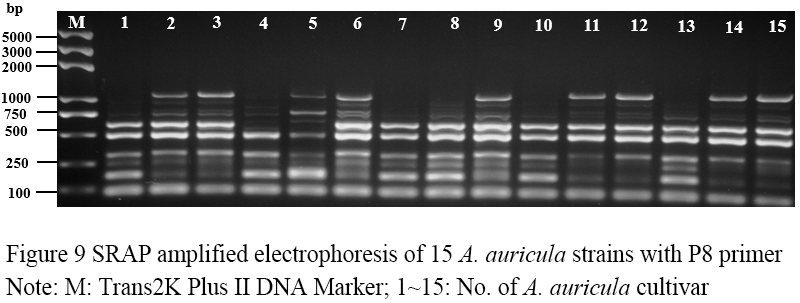 Figure 9 SRAP amplified electrophoresis of 15 A. auricula strains with P8 primer Note: M: Trans2K Plus DNA Marker; 1~15: No. of A. auricula cultivar |
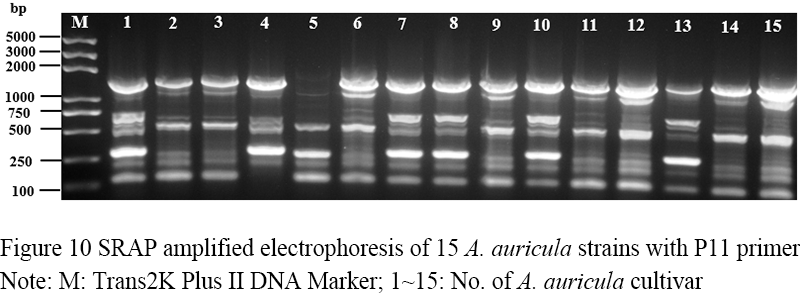 Figure 10 SRAP amplified electrophoresis of 15 A. auricula strains with P11 primer Note: M: Trans2K Plus DNA Marker; 1~15: No. of A. auricula cultivar |
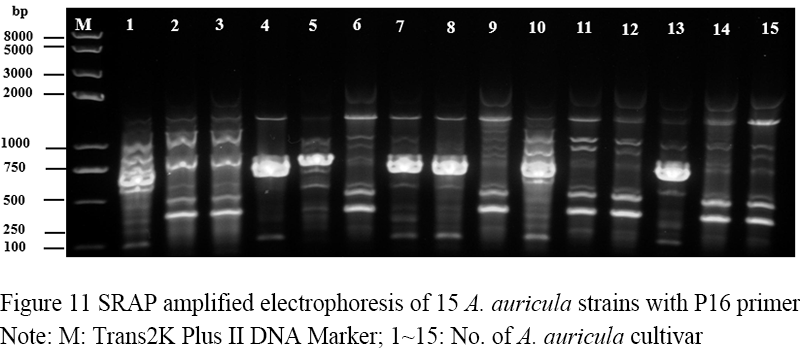 Figure 11 SRAP amplified electrophoresis of 15 A. auricula strains with P16 primer Note: M: Trans2K Plus II DNA Marker; 1~15: No. of A. auricula cultivar |
 Figure 12 SRAP amplified electrophoresis of 15 A. auricula strains with P17 primer Note: M: Trans2K Plus II DNA Marker; 1~15: No. of A. auricula cultivar |
The similarity coefficients of 15 A. auricula cultivars were calculated according to the electrophoresis band patterns of the amplified products, and the cluster analysis was performed using NTSYS software. It can be seen from the Phylogenetic tree (Figure 13) that the similarity coefficient of 15 A. auricula cultivars was 61%~100%, at around 76%. 15 A. auricula cultivars were divided into three categories, the first major categories as follows: 'Jiyou No. 1'; For the second categories: ‘Heiweibanjin’, 'Longjianghei+', 'Heishan', 'Gaochanwnag' and 'Jipin-8', 'Baolin 2', 'Diaodai', 'Heishan'; The third category is: 'Heimuer-2', 'Heimuer-1', 'Yante No.5', 'Hewei 15', 'Hei 29', 'Heimuer-2'. Among them, the similarity between 'Diaodai' and 'Heishan' strains was 95%, indicating that the genetic difference between them was very small. The similarity coefficient between 'Jiyou No.1' and other strains was 59%, indicating that the similarity was relatively small and the genetic relationship was far.
 Figure 13 Phylogenetic tree of 15 A. auricula strains based on SRAP |
2 Discussion
A.auricula is the second largest edible fungus in China, with an output of 62.7×108 kg in 2017, accounting for more than 99% of the world's total output. It is exported to Japan, Europe and America, earning an average annual foreign exchange of more than one billion dollars. At present, A. auricula has formed an industrial development trend of "A. auricula from North to South in China", with cultivated area increasing year by year (Yao, 2012). With the rapid expansion of the development scale of the A. auricula industry, the market is in urgent need of excellent A. auricula varieties to meet the needs of the development of the A. auricula industry. The germplasm resources of A. auricula are the most important basic production materials of A. auricula industry, which directly affect the yield and quality of A. auricula, as well as the premise and guarantee of variety breeding. At present, there are many cultivated species of A. auricula, and the phenomenon of fungus degradation and confusion is serious.
The flat-stand method is widely used in the classification and identification of edible fungi to determine the genetic relationship between strains through the antagonistic reaction between mycelia. When the contact part between mycelia shows the uplift type, ravine type, color linear type and isolated type, different strains are judged (Fang et al., 2012; Chen et al., 2014). In this study, the results of affinity test showed that antagonism of 15 A. auricula strains mostly showed two incompatibilities, i.e., non uplift type and ravine type. However, there was no antagonistic reaction between 5 pairs of A. auricula ('Diaodai' and 'Heimuer 2', 'Heiweibanjin' and 'Heilongjiang+', 'Longjianghei+' and 'Jipin-8', 'Heiweibanjin' and 'Jipin-8', 'Heiweibanjin' and 'Jipin 8', 'Heimuer-2' and 'Heimuer-1'), while 'Jiyou No.1' had obvious antagonistic reaction with other strains.
In recent years, DNA molecular marker technology has been widely used in the classification and identification of edible fungi, among which rDNA-ITS method is the most widely used method for bacterial species identification (Gou et al., 2017). In this study, 15 A. auricular cultivars were divided into 3 groups by rDNA-ITS method, among which 'Heimuer-1' and 'Heimuer-2', 'Heiwaibanjin' and 'Longjianghei+', 'Diaodai' and 'Heishan' had no genetic difference, but 'Jiyou No.1' had great genetic difference with other cultivars.
Yao et al. (2017) showed that SRAP method was good for genetic diversity analysis of A. Auricular. In this study, the polymorphic fragments amplified by SRAP primers accounted for 77.8% of the total number, indicating that the bands amplified by SRAP primers were rich in polymorphism and could be used for genetic relationship analysis. The 15 cultivated species of A. auricular were divided into 3 categories by cluster analysis, among which the similarity between 'Diaodai' and 'Heishan' strains was 95%, indicating that the genetic difference between them was very small, and the similarity between 'Jiyou No.1' and other strains was 59%, indicating that the similarity was relatively small and the genetic relationship was far.
In this study, 15 cultivated species of A. auricular were identified by strain affinity test, ITS sequence analysis and SRAP cluster analysis. The results showed that ITS and SRAP could well cluster A. auricular, and the and the results were similar. Combination of strain compatibility test can better applied to genetic diversity analysis of A. auricula. In addition, the identification results also showed that the polymorphism of A. auricular cultivars was low, and some genetic differences among different cultivars were small. This study clarified the genetic relationship of 15 cultivated species of A. auricula, which has certain guiding significance for the resource evaluation and genetic breeding research of A. Auricula.
3 Materials and Methods
3.1 Experimental material
The tested Trichoderma viride strains were provided by Laboratory of Bioengineering, Institute of Biology, Hebei Academy of Sciences. The tested A. auricula strains and their sources are as follows (Table 3).
PDA medium (1 L): potato 200 g, cut into small pieces, boil for 0.5 h, filter with gauze, add 20 g glucose and 15 g AGAR, pH is natural, divide into 500 mL triangle flask, autoclave sterilization at 115℃ for 30 min.
Liquid PDA medium (1 L): PDA without AGAR.
3.2 Compatibility test of A. auricular cultivated species
Use a 90 mm diameter petri dish, and 30 mL PDA medium was added into each dish quantitatively with peristaltic pump. 15 kinds of A. auricular were inserted into the center of the plate, respectively, and cultured at 25℃ for 7 days. With 5 mm punch up along one edge of the A. auricular colonies punch. Three different varieties of bacterial blocks were inserted into each dish and arranged in triangles, and cultured at 25°C for 15 days, respectively. Observed and recorded the hypha growth and antagonism. Among the strains, the ravine type, uplift type, color line type and isolation type were all incompatible, and they were determined as different strains.
3.3 DNA extraction of A. auricular strain
A 90 mm diameter petri dish was used, and 30 mL PDA medium was added to each dish with a quantitative peristaltic pump. After the medium was solidified, sterilized cellophane was placed on the surface. In the center of the medium, 15 kinds of A. auricular species with a diameter of 5 mm were added, and cultured in a constant temperature incubator at 25℃ for 10 days. Mycelia were collected. Total DNA was extracted from 15 species of A. auricular mycelia by Tiangen plant genome DNA extraction kit DP305 (Tiangen Biotech (Beijing) Co., Ltd.), and DNA was quantified by NanoDrop ND-2000 spectrophotometer.
3.4 ITS identification of A. auricular cultivar strain
The universal primers ITS1 (5'-TCCGTagGTGaACCTGCGG-3') and ITS4 (5'-TCCTCCGCTTattGATatGC-3') were used to amplify the DNA of A. Auricular. PCR reaction system: 25 µL 2×Easy Taq PCR Super Mix AS111 (TransGen Biotech Co., Ltd.), 1 µL of upstream and downstream primers, 2 µL of DNA template, supplemented with ddH2O to 50 µL. PCR amplification conditions: pre-denaturation at 94℃ for 2 min, denaturation at 94℃ for 40 s, renaturation at 52℃ for 1 min, extension at 72℃ for 1 min, a total of 35 cycles, holding at 72℃ for 5 min, storage at 4℃, PCR products were detected by 1% agarose gel electrophoresis (Liu et al., 2019). PCR purification kit EP101 (TransGen Biotech Co., Ltd.) was used to purify the amplification products, and KIT CT301 (TransGen Biotech Co., Ltd.) was used to connect the purified PCR products to T3 vector. The LB plate containing Amp was used to screen the blue and white spots, and the positive clones were selected. 50 positive clones were selected from each sample and sent to Sangon Biotech (Shanghai) Co., Ltd for sequencing. The sequences obtained were compared with the known sequences in NCBI database (BLAST). MEGA 6.0 software was used. Phylogenetic tree was constructed by proximity method (Tamura et al., 2013).
3.5 SRAP identification of cultivated species of A. Auricular
17 pairs of SRAP primers (Table 4) were selected to amplify the DNA of 15 A. auricular cultivars (Yao et al., 2017; Liu et al., 2018). PCR reaction system: 12.5 µL 2×Easy Taq PCR Super Mix AS111 (TransGen Biotech Co., Ltd.), 0.5 µL upstream and downstream primers and 1 µL DNA template were added respectively, and ddH2O was supplemented to 30 µL. PCR amplification conditions: pre-denaturation at 94℃ for 5 min, denaturation at 95℃ for 30 s, renaturation at 35℃ for 30 s, extension at 72℃ for 1 min, 5 cycles; After denaturation at 95℃ for 30 s, annealing at 50℃ for 1 min, extension at 72℃ for 1 min, 35 cycles, extension at 72℃ for 10 min, and preservation at 4℃, PCR products were detected by 1% agarose gel electrophoresis.
.png) Table 4 Sequences of primers used for SRAP analysis |
Authors’ contributions
LKA was the executor of the study, responsible for the experiment operation, results analysis and paper writing. MH and LM participated in the SRAP experiment and part of the analysis, ZGW and YSL participated in the collection of cultivated species of A. auricular, and LSS was responsible for guiding the experiment design and paper modification. All authors read and approved the final manuscript.
Acknowledgements
This study was supported by the Science and Technology Program of Hebei Academy of Sciences (19309) and the High-level Talents Training Program of Hebei Academy of Sciences (2020G05).
Chen Y., Yao F.J., Zhang Y.M., and Fang M., 2014, Study on breeding of A New A. auriculala-judae (Bull.) Quél. cultivar, Beifang Yuanyi (Northern Horticulture), 38(8): 133-134
Du P., Cui B.K., Zhang C.F., Cui B.K., Zhang C.F., and Dai Y.C., 2013, Genetic diversity of wild A. auriculala-judae revealed by ISSR analysis, Biochem. Syst. Ecol., 48: 199-205
https://doi.org/10.1016/j.bse.2012.11.011
Fang M., Yao F.J., Wang X.E., Chen Y., Liu G.J., and Zhang Y.M., 2012, Studies on breeding of a new A. auriculala cultivar, Junwu Yanjiu (Journal of Fungal Research), 10(4): 263-265
Gou F.C., Hou Y.L., Song B., Huang X.W., Song Z.Q., and Ding X., 2017, The application of rDNA-ITS sequence analysis method and traditional classification method in the classification of fungi, Xihua Shifan Daxue Xuebao (Journal of China West Normal University (Natural Sciences)), 38(4): 382-386
Jin H., Zou J.X., Song C.F., Li C.T., and Piao R.Z., 2012, Phylogeny tree analysis of Flammulina velutipes based on rDNA-ITS sequences, Zhongguo Shiyongjun (Edible Fungi of China), 31(4): 37-39, 50
Liu F.Y., Wu R.G., Liu X.R., Wang A.X., Wang Y.X., Deng W.M., and Zhao J.M., 2018, Agronomic traits and genetic characteristic of a new A. auriculala – judae variety, Nan'er No.1, Fujian Nongye Xuebao (Fujian Journal of Agricultural Sciences), 33(12): 1264-1269
Liu J., Ma C.Y., Xiang X.J., Li C., Bian Y.B., Zhang J., and Xiao Y., 2017, Constructing core collections of Chinese wild Lentinula edodes strains based on SRAP markers, Shiyongjun Xuebao (Acta Edulis Fungi), 24(1): 7-17
Liu K.A., Zhang G.W., Ma H., Yin S.L., Liu M., and Li S.S., 2019, Breeding and cultivation formula of Flammulina velutipes of anti-Trichoderma harzianum, Beifang Yuanyi (Northern Horticulture), 43(1): 150-157
Liu X.T., Guo J.F., Wang S.Y., Li Y.J., and Na R., 2015, Identification of several edible mushrooms in Inner Mongolia by rDNA-ITS, Shiyao Yongjun (Edible and Medicinal Mushrooms), 23(5): 301-306
Shen J.W., ed., 2014, Shiyongjun Shengchan Jishu Daquan
Shi Y.L., Liu B.W., Gu X.Y., Guo J.Y., Zheng S.Y., and Zhang Q.Q., 2019, Application of Biochemical and ISSR molecular markers in identification of A. auriculalar, Beifang Yuanyi (Northern Horticulture), 43(9): 136-141
Tamura K., Stecher G., Peterson D., Filipski A., and Kumar S., 2013, MEGA6: Molecular evolutionary genetics analysis version 6.0, Mol. Biol. Evol., 30(12): 2725-2729
https://doi.org/10.1093/molbev/mst197
PMid:24132122 PMCid:PMC3840312
Wang D.D., Lin C.Y., Ma X.B., and Liu Y., 2018, Establishment and optimization of SRAP reaction system for wild relatives of garlic, Fenzi Zhiwu Yuzhong (Molecular Plant Breeding), 16(23): 7731-7739
Wang H., Niu Y.R., Rong C.B., Liu Y., Wang S.X., and Song S., 2019, Genetic diversity analysis of 19 Lentinus edodes strains based on ISSR, SRAP and TRAP markers, Jiangsu Nongye Kexue (Jiangsu Agricultural Sciences), 47(17): 54-59
Wang H.X., and Liu Q.H., eds., 2014, Mushroom cultivation, 2st edition, China Agricultural University Press, Beijing, China, pp.
Wang Y., Wan L.C., Huang C.Y., Han J.D., Li J., Yang P., Yao Q., Gong Z.Y., and Xie H.Y., 2019, Genetic diversity of Grifola frondosa based on integrated ISSR and SRAP analysis, Shiyongjun Xuebao (Acta Edulis Fungi), 26(3): 26-36
Yao F.J., Lu L.X., Wang P., Zhang Y.M., Yao Y.W., and Li H.Y., 2017, Verification of genuineness for spawn of Auricularia heimuer by SRAP, Beifang Yuanyi (Northern Horticulture), 41(24): 178-182
Yao F.J., 2012, The cultivation spread of A. auriculala from north to south in China, Zhongguo Shiyongjun (Edible Fungi of China), 31(1): 61-62
Zhang X., Li Q., Liu H.J., Liu K.R., Zhang Y.X., Liu T.F., and Yang Y.D., 2013, Application of SRAP and ITS molecular markers in genetic diversity of wild A. auriculala in Daxing'anling area, Heilongjiang Nongye Kexue (Heilongjiang Science), (4): 18-21
Zhu J.X., Li L.J., Zhao S.F., Chen H., and Jiang S.X., 2018, Genetic diversity and phylogenetic relationship analysis of 23 Stropharia rugosoannulata strains based on SRAP molecular marker, Shandong Nongye Kexue (Shandong Agricultural Sciences), 50(3): 22-28



. PDF(0KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Kunang Liu
. Hong Ma
. Meng Liu
. Genwei Zhang
. Shuli Yin
. Shusheng Li
Related articles
. A. auricula
. rDNA-ITS
. SRAP
. Genetic diversity
Tools
. Email to a friend
. Post a comment